


Next: The Statistical Model
Up: Constructing Physical Maps from
Previous: Constructing Physical Maps from
Physical mapping using hybridization fingerprints of short
oligonucleotides was first suggested by Poustka et al. in 1986
[7]. In this technique short labeled DNA sequences, or
probes, attach, or hybridize, to positions along the target DNA
matching their own DNA sequence. The probes are nonunique, i.e.,
they occur at many points along the genome, and typically
hybridize with
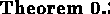 of the clones. Overlapping clones can
be identified by their similar fingerprints. See figure
9.9 for an illustration of this
hybridization scenario. [7] suggested this method in order
to eliminate the need to process individual clones in the
restriction digestion technique. They reported preliminary
computer simulations demonstrating feasibility, and suggested the
use of Bayesian inference in data analysis. More detailed
strategies were offered by Michiels et al. [6]. A
likelihood ratio based on a detailed statistical model was used to
make overlap decisions, and a discussion of experimental errors
was also included. Craig et al. [2] used short
oligonucleotides in the ordering of cosmid clones covering the
Herpes Simplex Virus (HSVI) genome. The clones were ordered
manually. As each probe occurred only once or twice along the
short (
of the clones. Overlapping clones can
be identified by their similar fingerprints. See figure
9.9 for an illustration of this
hybridization scenario. [7] suggested this method in order
to eliminate the need to process individual clones in the
restriction digestion technique. They reported preliminary
computer simulations demonstrating feasibility, and suggested the
use of Bayesian inference in data analysis. More detailed
strategies were offered by Michiels et al. [6]. A
likelihood ratio based on a detailed statistical model was used to
make overlap decisions, and a discussion of experimental errors
was also included. Craig et al. [2] used short
oligonucleotides in the ordering of cosmid clones covering the
Herpes Simplex Virus (HSVI) genome. The clones were ordered
manually. As each probe occurred only once or twice along the
short ( 140KB) genome, this experiment does not represent
the general problem.
140KB) genome, this experiment does not represent
the general problem.
Figure:
Clones and
non-unique probes. The clones are the horizontal lines. The random
occurrences of a single nonunique probe are marked by the dotted
vertical lines.
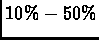 |
Figure 9.9:
Physical map
example. The short horizontal lines are the clones with their x
coordinates corresponding to the position on the target genome.
The y coordinates correspond to the clone order in the constructed
map. Note that each point on the target genome is covered by many
clones. The total length of the clones divided by the length of
the genome is called the clone coverage (10 in this example).
 |
The location of the clones along the target genome is not directly
known to the experimenters. Mapping data (such as hybridization
data) produced by the experiment is used to reconstruct the map. A
list assigning every clone its estimated position along the genome
is a solution to the mapping problem. According to equation
9.7 in theorem
9.3, with sufficient coverage the whole
map is usually one contig. A plot of clone order in the
constructed map vs. real clone position (see figure
9.9) provides a visual measure for map
quality. If the order of the clones in the constructed map is
completely correct then the computed left endpoints of clones
increase as their true value increase (or decrease, if the orders
in the true and constructed maps happen to be reversed, as in the
examples of figures
9.8,9.9).
Minor ordering errors are seen as small deviations from the
monotonicity, as in figure 9.8, show
the construction is still essentially correct. Very small errors,
which do not change the clone order, cannot be observed from the
plot. A completely random solution will correspond to randomly
placed clones, whereas a nonrandom solution containing several
large errors will translate into several randomly placed broken
contigs with an approximately correct intra - contig order.
Next: The Statistical Model
Up: Constructing Physical Maps from
Previous: Constructing Physical Maps from
Peer Itsik
2001-01-09