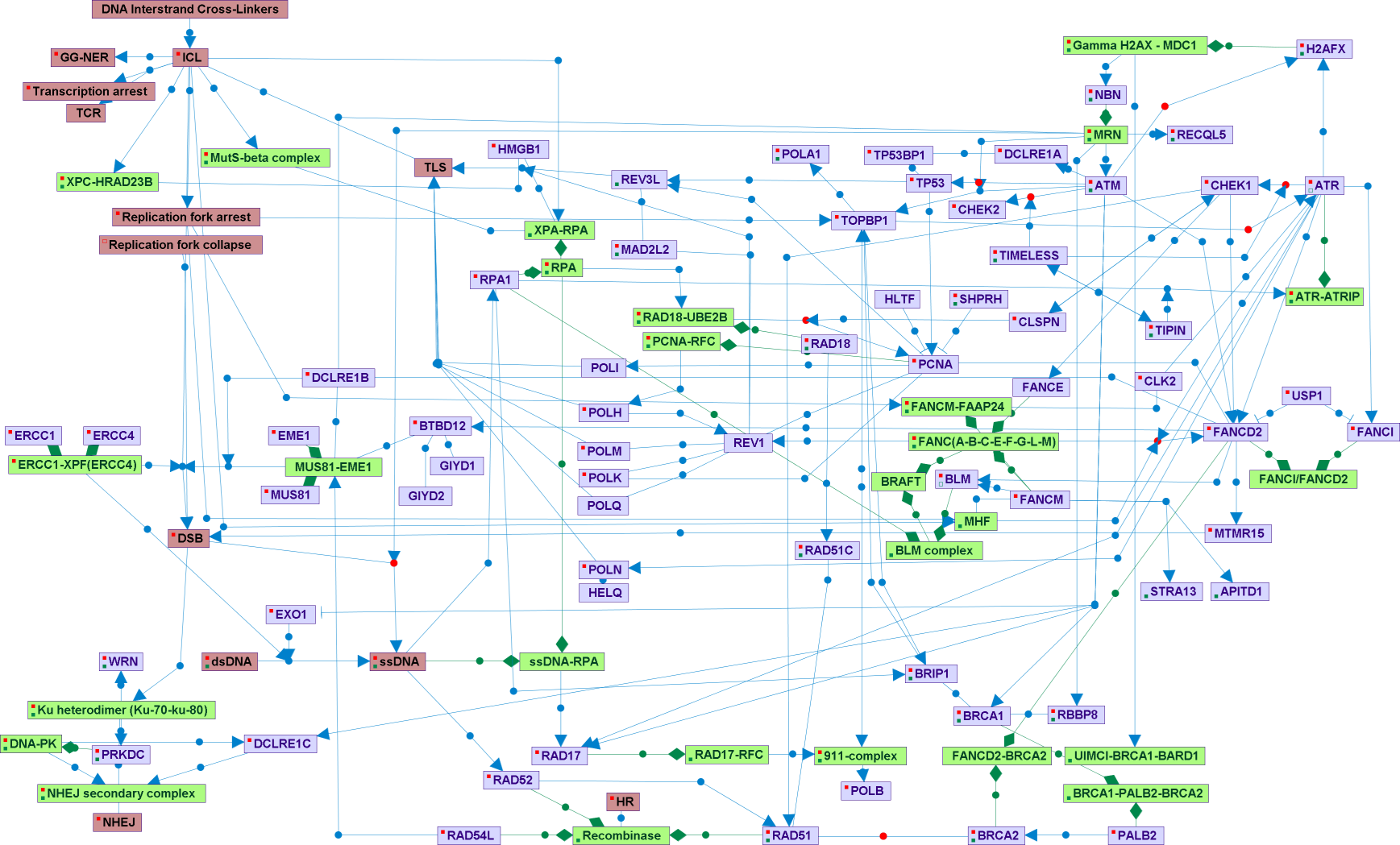
DNA Interstrand crosslinks (ICLs) prevent DNA strand separation and can cause absolute blocks to DNA replication and/or DNA transcription. Natural agents like nitrous acid, malondialdehyde (a product of lipid peroxidation), and other bifunctional alkylating agents can cause ICLs. Platinum compounds and psoralen are used as ICL-forming compounds in cancer therapy. The major ICL-repair pathway in mammals depends on homologous recombination. ICLs represent an impenetrable block for replication during the S-phase, causing replication forks arrest and collapse. Adjacent to the collapsed replication forks double strand breaks (DSB) occurred. Then, there is a release of the ICL attachment to one of the DNA strands, following by homologous recombination (HR). The NER proteins ERCC1 and ERCC4 form a nuclease complex essential to the incision step (DSB formation). The MUSS81-EME1complex has a similar role. The mismatch repair (MMR) proteins that forms the MUTS-beta complex, and additional NER proteins (XPA and XPC) contributes to the ICLs damage resolution, beyond their canonic roles in MMR and NER. There is also a minor, recombination-independent repair pathway. Nucleotide-excision repair (NER) genes are involved in this pathway, and some error-prone DNA polymerases, that enable translesion synthesis (TLS). PCNA ubiquitination is the TLS-activating signal, and REV3L and REV1 are the main components of this branch. The Fanconi proteins are essential for both HR and TLS pathways. The Ubiquitination of FANCD2 is the key signaling event, although ATM, ATR, and CHEK1 phosphorylates FANCD2 and other Fanconi proteins. Additional important proteins in the contest of damage recognition, repair and signaling are TOPBP1, BRCA1, BLM, TIMELESS, CLSPN, and TIPIN.